Medisch expert van het artikel

Nieuwe publicaties
hypofyse
Laatst beoordeeld: 04.07.2025

Alle iLive-inhoud wordt medisch beoordeeld of gecontroleerd op feiten om zo veel mogelijk feitelijke nauwkeurigheid te waarborgen.
We hebben strikte richtlijnen voor sourcing en koppelen alleen aan gerenommeerde mediasites, academische onderzoeksinstellingen en, waar mogelijk, medisch getoetste onderzoeken. Merk op dat de nummers tussen haakjes ([1], [2], etc.) klikbare links naar deze studies zijn.
Als u van mening bent dat onze inhoud onjuist, verouderd of anderszins twijfelachtig is, selecteert u deze en drukt u op Ctrl + Enter.
De hypofyse (hypofyse, s.glandula pituitaria) bevindt zich in de hypofysaire fossa van de sella turcica van het wiggenbeen en is van de schedelholte gescheiden door een uitloper van de dura mater van de hersenen, die het diafragma van de sella vormt. Via de opening in dit diafragma is de hypofyse verbonden met het infundibulum van de hypothalamus van het diencephalon. De dwarsdoorsnede van de hypofyse is 10-17 mm, de anteroposterieure doorsnede is 5-15 mm, de verticale doorsnede is 5-10 mm. De massa van de hypofyse bij mannen is ongeveer 0,5 g, bij vrouwen 0,6 g. De hypofyse is uitwendig bedekt met een kapsel.
In overeenstemming met de ontwikkeling van de hypofyse vanuit twee verschillende rudimenten, worden er twee lobben in het orgaan onderscheiden - de voorste en de achterste. De adenohypofyse, of voorkwab (adenohypofyse, s.lobus anterior), is groter en maakt 70-80% uit van de totale massa van de hypofyse. Deze is dichter dan de achterste kwab. In de voorkwab wordt een distaal deel (pars distalis) onderscheiden, dat het voorste deel van de hypofysefossa beslaat, een intermediair deel (pars intermedia), gelegen op de grens met de achterste kwab, en een tubereus deel (pars tuberalis), dat naar boven loopt en aansluit op het infundibulum van de hypothalamus. Vanwege de overvloed aan bloedvaten heeft de voorkwab een lichtgele kleur met een roodachtige tint. Het parenchym van de hypofysevoorkwab bestaat uit verschillende soorten kliercellen, met daartussen sinusvormige bloedcapillairen. De helft (50%) van de adenohypofysecellen zijn chromofiele adenocyten, met fijnkorrelige granula in hun cytoplasma die goed kleuren met chroomzouten. Dit zijn acidofiele adenocyten (40% van alle adenohypofysecellen) en basofiele adenocyten (10%). Basofiele adenocyten omvatten gonadotrope, corticotrope en thyrotrope endocrinocyten. Chromofobe adenocyten zijn klein, hebben een grote celkern en een kleine hoeveelheid cytoplasma. Deze cellen worden beschouwd als voorlopers van chromofiele adenocyten. De overige 50% van de adenohypofysecellen zijn chromofobe adenocyten.
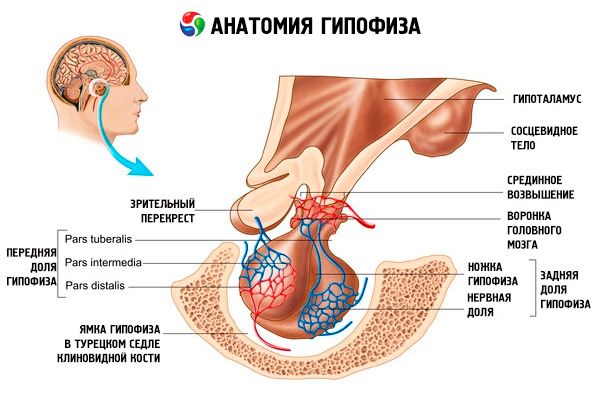
De neurohypofyse, of achterkwab (neurohypofyse, s.lobus posterior), bestaat uit de neurale kwab (lobus nervosus), gelegen in het achterste deel van de hypofyse, en de trechter (infundibulum), gelegen achter het tubereuze deel van de adenohypofyse. De achterkwab van de hypofyse wordt gevormd door neurogliacellen (hypofysecellen), zenuwvezels die van de neurosecretoire kernen van de hypothalamus naar de neurohypofyse lopen, en neurosecretoire bloedlichaampjes.
De hypofyse is via zenuwvezels en bloedvaten functioneel verbonden met de hypothalamus van het diencephalon, die de activiteit van de hypofyse reguleert. De hypofyse en de hypothalamus, samen met hun neuro-endocriene, vasculaire en zenuwverbindingen, worden doorgaans beschouwd als het hypothalamus-hypofysestelsel.
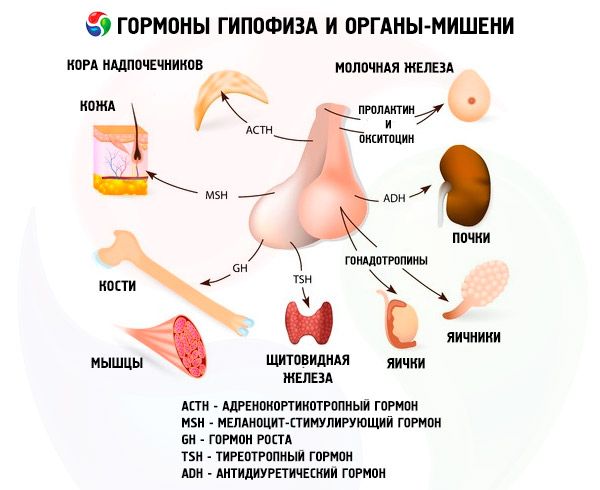
Hormonen van de hypofysevoorkwab en de hypofyseachterkwab beïnvloeden vele lichaamsfuncties, voornamelijk via andere endocriene klieren. In de hypofyse produceren acidofiele adenocyten (alfacellen) somatotroop hormoon (groeihormoon), dat deelneemt aan de regulering van groei- en ontwikkelingsprocessen bij jonge organismen. Corticotrope endocrinocyten scheiden adrenocorticotroop hormoon (ACTH) af, dat de afgifte van steroïde hormonen door de bijnieren stimuleert. Thyrotrope endocrinocyten scheiden thyrotroop hormoon (TSH) af, dat de ontwikkeling van de schildklier beïnvloedt en de productie van schildklierhormonen activeert. Gonadotrope hormonen: follikelstimulerend hormoon (FSH), luteïniserend hormoon (LH) en prolactine - beïnvloeden de seksuele rijping van het lichaam en reguleren en stimuleren de ontwikkeling van follikels in de eierstokken, de ovulatie, de groei van de melkklieren en de melkproductie bij vrouwen, en het proces van spermatogenese bij mannen. Deze hormonen worden geproduceerd door basofiele adenocyten (bètacellen). Lipotrope factoren van de hypofyse worden hier ook afgescheiden, en beïnvloeden de mobilisatie en het gebruik van vetten in het lichaam. In het tussenliggende deel van de voorkwab wordt melanocytstimulerend hormoon gevormd, dat de vorming van pigmenten - melaninen - in het lichaam reguleert.
Neurosecretoire cellen van de supraoptische en paraventriculaire kernen in de hypothalamus produceren vasopressine en oxytocine. Deze hormonen worden via axonen, die de hypothalamus-hypofysebaan vormen, naar de cellen van de hypofyseachterkwab getransporteerd. Vanuit de hypofyseachterkwab komen deze stoffen in het bloed terecht. Het hormoon vasopressine heeft een vasoconstrictieve en antidiuretische werking, waardoor het ook wel antidiuretisch hormoon (ADH) wordt genoemd. Oxytocine stimuleert de contractiliteit van de baarmoederspieren, verhoogt de melkproductie van de melkklier, remt de ontwikkeling en functie van het corpus luteum en beïnvloedt veranderingen in de tonus van de gladde (niet-dwarsgestreepte) spieren van het maag-darmkanaal.
Ontwikkeling van de hypofyse
De voorkwab van de hypofyse ontwikkelt zich vanuit het epitheel van de achterwand van de orale baai in de vorm van een ringvormige uitgroei (het zakje van Rathke). Deze ectodermale uitstulping groeit naar de bodem van het toekomstige derde ventrikel. Daarheen, vanaf de onderkant van het tweede hersenblaasje (de toekomstige bodem van het derde ventrikel), groeit een uitloper, waaruit zich de grijze knobbel van het infundibulum en de achterkwab van de hypofyse ontwikkelen.
Vaten en zenuwen van de hypofyse
Vanuit de arteria carotis interna en de bloedvaten van de grote hersenen lopen de arteria hypofyse superior en inferior naar de hypofyse. De arteria hypofyse superior lopen naar de nucleus grayes en het infundibulum van de hypothalamus, anastomoseren hier met elkaar en vormen haarvaten die het hersenweefsel binnendringen - het primaire hemocapillair netwerk. Vanuit de lange en korte lussen van dit netwerk worden de poortaders gevormd, die naar de voorkwab van de hypofyse lopen. In het parenchym van de voorkwab van de hypofyse splitsen deze aders zich op in brede sinusvormige haarvaten, die het secundaire hemocapillair netwerk vormen. De achterkwab van de hypofyse wordt voornamelijk van bloed voorzien door de arteria hypofyse inferior. Er zijn lange arteriële anastomosen tussen de arteria hypofyse superior en inferior. De uitstroom van veneus bloed uit het secundaire hemocapillaire netwerk vindt plaats via een systeem van aderen die uitmonden in de caverneuze en intercaverneuze sinussen van de dura mater van de hersenen.
Sympathische zenuwvezels die samen met slagaders het orgaan binnenkomen, spelen een rol bij de innervatie van de hypofyse. Postganglionaire sympathische zenuwvezels vertrekken vanuit de plexus van de arteria carotis interna. Daarnaast bevinden zich in de achterkwab van de hypofyse talrijke uiteinden van de uitlopers van neurosecretoire cellen, gelegen in de kernen van de hypothalamus.
Leeftijdsgebonden kenmerken van de hypofyse
Het gemiddelde gewicht van de hypofyse bij pasgeborenen bedraagt 0,12 gram. Het gewicht van het orgaan verdubbelt rond de leeftijd van 10 jaar en verdrievoudigt rond de leeftijd van 15 jaar. Rond de leeftijd van 20 jaar bereikt het gewicht van de hypofyse zijn maximum (530-560 mg) en blijft het vrijwel onveranderd in de daaropvolgende levensfasen. Na 60 jaar wordt een lichte afname in het gewicht van deze endocriene klier waargenomen.
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
Hypofysehormonen
De eenheid van de zenuw- en hormoonregulatie in het lichaam wordt gewaarborgd door de nauwe anatomische en functionele verbinding tussen de hypofyse en de hypothalamus. Dit complex bepaalt de toestand en werking van het gehele endocriene systeem.
De belangrijkste endocriene klier die een aantal peptidehormonen produceert die de functie van de perifere klieren rechtstreeks reguleren, is de hypofyse. Het is een roodgrijze boonvormige formatie bedekt met een fibreuze capsule met een gewicht van 0,5-0,6 g. Deze varieert enigszins afhankelijk van het geslacht en de leeftijd van een persoon. De algemeen aanvaarde verdeling van de hypofyse in twee lobben die verschillen in ontwikkeling, structuur en functie blijft: de voorste distale - adenohypofyse en de achterste - neurohypofyse. De eerste maakt ongeveer 70% uit van de totale massa van de klier en wordt conventioneel verdeeld in het distale, intrechtervormige en intermediaire deel, de tweede - in het achterste deel, of lob, en de hypofysesteel. De klier bevindt zich in de hypofysefossa van de sella turcica van het wiggenbeen en is via de steel verbonden met de hersenen. Het bovenste deel van de voorkwab wordt bedekt door het chiasma opticum en de tractus opticum. De hypofyse wordt zeer rijk van bloed voorzien via aftakkingen van de arteria carotis interna (de arteria hypofyse superior en arteria inferior), evenals via aftakkingen van de cerebrum arteriae arteriae hypofyse. De arteria hypofyse superior en de arteria hypofyse inferior, de neurohypofyse, maken deel uit van de bloedvoorziening van de adenohypofyse en de arteria hypofyse inferior, en staan in contact met de neurosecretoire uiteinden van de axonen van de grote celkernen van de hypothalamus. De eerstgenoemde komen de eminentie medianus van de hypothalamus binnen, waar ze zich verspreiden in een capillair netwerk (de primaire capillaire plexus). Deze haarvaten (waarmee de uiteinden van de axonen van de kleine neurosecretoire cellen van de mediobasale hypothalamus in contact komen) verzamelen zich in de poortaderen die langs de hypofysesteel afdalen naar het parenchym van de adenohypofyse, waar ze zich weer splitsen in een netwerk van sinusvormige haarvaten (de secundaire capillaire plexus). Zo komt het bloed, dat eerst de mediane eminentie van de hypothalamus is gepasseerd, waar het verrijkt wordt met hypothalamische adenohypofysotrope hormonen (releasing hormones), de adenohypofyse binnen.
De uitstroom van met adenohypofysehormonen verzadigd bloed vanuit talrijke haarvaten van de secundaire plexus vindt plaats via het veneuze systeem, dat op zijn beurt uitmondt in de veneuze sinussen van de dura mater en vervolgens in de algemene bloedbaan. Het portaalsysteem van de hypofyse, met een afdalende bloedstroom vanuit de hypothalamus, is dus een morfofunctioneel onderdeel van het complexe mechanisme van neurohumorale controle van de tropische functies van de adenohypofyse.
De hypofyse wordt geïnnerveerd door sympathische vezels die de hypofysaire slagaders volgen. Deze vezels ontspringen uit postganglionaire vezels die door de plexus carotis interna lopen en verbonden zijn met de bovenste cervicale ganglia. De adenohypofyse wordt niet rechtstreeks geïnnerveerd vanuit de hypothalamus. De achterste kwab ontvangt zenuwvezels vanuit de neurosecretoire kernen van de hypothalamus.
De adenohypofyse is een zeer complexe formatie in zijn histologische architectuur. Hij heeft twee soorten kliercellen: chromofobe en chromofiele. Deze laatste worden op hun beurt onderverdeeld in acidofiele en basofiele (een gedetailleerde histologische beschrijving van de hypofyse is te vinden in het betreffende hoofdstuk van de handleiding). Er dient echter te worden opgemerkt dat de hormonen die worden geproduceerd door de kliercellen die het parenchym van de adenohypofyse vormen, vanwege de diversiteit van deze laatste, enigszins verschillen in hun chemische aard, en dat de fijnstructuur van de secreterende cellen moet overeenkomen met de biosynthese-eigenschappen van elk van hen. Soms worden echter overgangsvormen van kliercellen waargenomen in de adenohypofyse die in staat zijn om meerdere hormonen te produceren. Er zijn aanwijzingen dat het type kliercellen van de adenohypofyse niet altijd genetisch bepaald is.
Onder het diafragma van de sella turcica bevindt zich het trechtervormige deel van de voorkwab. Het omsluit de hypofysesteel en staat in contact met het grijze knobbeltje. Dit deel van de adenohypofyse wordt gekenmerkt door de aanwezigheid van epitheelcellen en een overvloedige bloedtoevoer. Het is ook hormonaal actief.
Het tussenliggende (middelste) deel van de hypofyse bestaat uit verschillende lagen van grote secretoire-actieve basofiele cellen.
De hypofyse vervult verschillende functies via haar hormonen. De voorkwab produceert adrenocorticotrope (ACTH), schildklierstimulerende (TSH), follikelstimulerende (FSH), luteïniserende (LH), lipotrope hormonen, evenals groeihormoon-somatotrope (STO) en prolactine. In de tussenkwab wordt melanocytstimulerend hormoon (MSH) gesynthetiseerd en in de achterkwab accumuleren vasopressine en oxytocine.
ACTH
Hypofysehormonen zijn een groep eiwit- en peptidehormonen en glycoproteïnen. Van de hormonen van de hypofyse is ACTH het best bestudeerd. Het wordt geproduceerd door basofiele cellen. De belangrijkste fysiologische functie is het stimuleren van de biosynthese en secretie van steroïde hormonen door de bijnierschors. ACTH vertoont ook melanocytstimulerende en lipotrope activiteit. In 1953 werd het in zuivere vorm geïsoleerd. Later werd de chemische structuur vastgesteld, bestaande uit 39 aminozuren bij mensen en een aantal zoogdieren. ACTH heeft geen soortspecificiteit. Momenteel vindt chemische synthese plaats van zowel het hormoon zelf als verschillende fragmenten van het molecuul, die actiever zijn dan natuurlijke hormonen. De structuur van het hormoon bestaat uit twee delen van de peptideketen, waarvan er één zorgt voor de detectie en binding van ACTH aan de receptor, en de andere voor een biologisch effect. Het bindt zich blijkbaar aan de ACTH-receptor door de interactie van de elektrische ladingen van het hormoon en de receptor. De rol van biologische effector van ACTH wordt vervuld door het fragment van het 4-10-molecuul (Met-Glu-His-Phen-Arg-Tri-Tri).
De melanocytstimulerende activiteit van ACTH is te danken aan de aanwezigheid van de N-terminale regio in het molecuul, bestaande uit 13 aminozuren en met een repeterende structuur van alfa-melanocytstimulerend hormoon. Deze regio bevat ook een heptapeptide, aanwezig in andere hypofysehormonen, met een adrenocorticotrope, melanocytstimulerende en lipotrope werking.
Het sleutelmoment in de werking van ACTH wordt beschouwd als de activering van het enzym proteïnekinase in het cytoplasma, met medewerking van cAMP. Gefosforyleerde proteïnekinase activeert het enzym esterase, dat cholesterolesters omzet in een vrije stof in vetdruppels. Het eiwit dat in het cytoplasma wordt gesynthetiseerd door fosforylering van ribosomen, stimuleert de binding van vrije cholesterol aan cytochroom P-450 en de overdracht ervan vanuit lipidedruppels naar mitochondriën, waar alle enzymen aanwezig zijn die zorgen voor de omzetting van cholesterol in corticosteroïden.
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
Schildklierstimulerend hormoon
TSH - thyrotropine - de belangrijkste regulator van de ontwikkeling en werking van de schildklier, de processen van synthese en secretie van schildklierhormonen. Dit complexe eiwit - glycoproteïne - bestaat uit alfa- en bèta-subeenheden. De structuur van de eerste subeenheid komt overeen met de alfa-subeenheid van luteïniserend hormoon. Bovendien komt deze grotendeels overeen bij verschillende diersoorten. De aminozuursequentie in de bèta-subeenheid van menselijk TSH is ontrafeld en bestaat uit 119 aminozuren. Opgemerkt kan worden dat de bèta-subeenheden van menselijk en rund-TSH in veel opzichten vergelijkbaar zijn. De biologische eigenschappen en de aard van de biologische activiteit van glycoproteïnehormonen worden bepaald door de bèta-subeenheid. Deze zorgt ook voor de interactie van het hormoon met receptoren in verschillende doelorganen. De bèta-subeenheid vertoont bij de meeste dieren echter pas specifieke activiteit nadat deze zich verbindt met de alfa-subeenheid, die als een soort activator van het hormoon fungeert. Laatstgenoemde induceert met gelijke waarschijnlijkheid luteïniserende, follikelstimulerende en thyrotrope activiteiten, bepaald door de eigenschappen van de bèta-subeenheid. De gevonden gelijkenis stelt ons in staat te concluderen dat deze hormonen in het evolutieproces zijn ontstaan uit één gemeenschappelijke voorouder; de bèta-subeenheid bepaalt ook de immunologische eigenschappen van de hormonen. Er wordt aangenomen dat de alfa-subeenheid de bèta-subeenheid beschermt tegen de werking van proteolytische enzymen en tevens het transport ervan van de hypofyse naar de perifere doelorganen faciliteert.
Gonadotrope hormonen
Gonadotropinen zijn in het lichaam aanwezig in de vorm van LH en FSH. De functionele functie van deze hormonen is over het algemeen beperkt tot het bevorderen van voortplantingsprocessen bij individuen van beide geslachten. Net als TSH zijn het complexe eiwitten - glycoproteïnen. FSH induceert de rijping van follikels in de eierstokken bij vrouwen en stimuleert de spermatogenese bij mannen. LH veroorzaakt follikelruptuur bij vrouwen met de vorming van het corpus luteum en stimuleert de secretie van oestrogenen en progesteron. Bij mannen versnelt hetzelfde hormoon de ontwikkeling van interstitieel weefsel en de secretie van androgenen. De effecten van gonadotropinen zijn van elkaar afhankelijk en treden synchroon op.
De dynamiek van de gonadotropinesecretie bij vrouwen verandert tijdens de menstruatiecyclus en is voldoende gedetailleerd bestudeerd. In de pre-ovulatoire (folliculaire) fase van de cyclus is het LH-gehalte vrij laag, terwijl het FSH-gehalte verhoogd is. Naarmate de follikel rijpt, neemt de oestradiolsecretie toe, wat een verhoogde productie van gonadotropines door de hypofyse en het optreden van zowel LH- als FSH-cycli bevordert. Geslachtshormonen stimuleren de secretie van gonadotropines.
De structuur van LH is momenteel vastgesteld. Net als TSH bestaat het uit twee subeenheden: a en b. De structuur van de alfa-subeenheid van LH bij verschillende diersoorten komt grotendeels overeen met de structuur van de alfa-subeenheid van TSH.
De structuur van de bèta-subeenheid van LH verschilt aanzienlijk van die van de bèta-subeenheid van TSH, hoewel deze vier identieke delen van de peptideketen heeft, bestaande uit 4-5 aminozuren. In TSH zijn deze gelokaliseerd op de posities 27-31, 51-54, 65-68 en 78-83. Aangezien de bèta-subeenheid van LH en TSH de specifieke biologische activiteit van de hormonen bepaalt, kan worden aangenomen dat homologe delen in de structuur van LH en TSH de verbinding van bèta-subeenheden met de alfa-subeenheid garanderen, en dat delen met een verschillende structuur verantwoordelijk zijn voor de specificiteit van de biologische activiteit van de hormonen.
Natuurlijk LH is zeer stabiel ten opzichte van de werking van proteolytische enzymen, maar de bèta-subeenheid wordt snel gesplitst door chymotrypsine. Deze subeenheid is bovendien moeilijk te hydrolyseren door het enzym. Het speelt een beschermende rol doordat het chymotrypsine geen toegang heeft tot de peptidebindingen.
Wat de chemische structuur van FSH betreft, hebben onderzoekers nog geen definitieve resultaten behaald. Net als LH bestaat FSH uit twee subeenheden, maar de bèta-subeenheid van FSH verschilt van de bèta-subeenheid van LH.
Prolactine
Een ander hormoon, prolactine (lactogeen hormoon), speelt een actieve rol in voortplantingsprocessen. De belangrijkste fysiologische eigenschappen van prolactine bij zoogdieren manifesteren zich in de vorm van stimulatie van de ontwikkeling van de melkklieren en de lactatie, en de groei van de talgklieren en inwendige organen. Het bevordert de manifestatie van het effect van steroïden op secundaire geslachtskenmerken bij mannetjes, stimuleert de secretoire activiteit van het corpus luteum bij muizen en ratten en neemt deel aan de regulering van de vetstofwisseling. Er is de laatste jaren veel aandacht besteed aan prolactine als regulator van moederlijk gedrag; deze polyfunctionaliteit wordt verklaard door de evolutionaire ontwikkeling ervan. Het is een van de oudste hypofysehormonen en wordt zelfs bij amfibieën aangetroffen. Momenteel is de structuur van prolactine bij sommige zoogdiersoorten volledig ontrafeld. Tot voor kort uitten wetenschappers echter twijfels over het bestaan van een dergelijk hormoon bij mensen. Velen geloofden dat de functie ervan werd vervuld door het groeihormoon. Er is nu overtuigend bewijs voor de aanwezigheid van prolactine bij mensen en de structuur ervan is gedeeltelijk ontrafeld. Prolactinereceptoren binden actief groeihormoon en placentair lactogeen, wat wijst op een gemeenschappelijk werkingsmechanisme van de drie hormonen.
Somatotropine
Het groeihormoon somatotropine heeft een nog breder werkingsspectrum dan prolactine. Net als prolactine wordt het geproduceerd door acidofiele cellen van de adenohypofyse. STH stimuleert de skeletgroei, activeert de eiwitsynthese, heeft een vetmobiliserende werking en bevordert een toename van de lichaamsgrootte. Bovendien coördineert het stofwisselingsprocessen.
De deelname van het hormoon aan dit laatste wordt bevestigd door het feit dat er een sterke toename is van de afscheiding ervan door de hypofyse, bijvoorbeeld wanneer het bloedsuikergehalte daalt.
De chemische structuur van dit menselijke hormoon is nu volledig vastgesteld: 191 aminozuren. De primaire structuur is vergelijkbaar met die van chorion-somatomammotropine of placentair lactogeen. Deze gegevens wijzen op een significante evolutionaire verwantschap tussen de twee hormonen, hoewel ze verschillen in biologische activiteit.
De hoge soortspecificiteit van het betreffende hormoon moet worden benadrukt. Zo is STH van dierlijke oorsprong inactief bij mensen. Dit wordt verklaard door zowel de reactie tussen de receptoren van STH bij mensen en dieren als de structuur van het hormoon zelf. Momenteel wordt onderzoek gedaan naar de identificatie van actieve centra in de complexe structuur van STH die biologische activiteit vertonen. Individuele fragmenten van het molecuul met andere eigenschappen worden bestudeerd. Zo werd na hydrolyse van humaan STH door pepsine een peptide geïsoleerd, bestaande uit 14 aminozuren en overeenkomend met de molecuulsecties 31-44. Het had geen groei-effect, maar overtrof het natuurlijke hormoon aanzienlijk in lipotrope activiteit. Menselijk groeihormoon heeft, in tegenstelling tot het vergelijkbare hormoon van dieren, een significante lactogene activiteit.
De adenohypofyse synthetiseert veel peptide- en eiwitstoffen met een vetmobiliserende werking, en de hypofysehormonen – ACTH, STH, TSH en andere – hebben een lipotrope werking. De laatste jaren is er vooral aandacht besteed aan bèta- en y-lipotrope hormonen (LPG). De biologische eigenschappen van bèta-LPG zijn zeer gedetailleerd bestudeerd; naast lipotrope activiteit heeft het ook een melanocytstimulerend, corticotropinestimulerend en hypocalciëmisch effect, en produceert het een insuline-achtig effect.
Momenteel is de primaire structuur van schapen-LPG (90 aminozuren) en lipotrope hormonen van varkens en runderen ontrafeld. Dit hormoon is soortspecifiek, hoewel de structuur van het centrale deel van bèta-LPG bij verschillende soorten hetzelfde is. Het bepaalt de biologische eigenschappen van het hormoon. Een van de fragmenten van dit deel bevindt zich in de structuur van alfa-MSH, bèta-MSH, ACTH en bèta-LPG. Er wordt verondersteld dat deze hormonen tijdens de evolutie uit dezelfde voorloper zijn ontstaan. γ-LPG heeft een zwakkere lipotrope activiteit dan bèta-LPG.
Melanocytstimulerend hormoon
Dit hormoon, gesynthetiseerd in de tussenkwab van de hypofyse, stimuleert de biosynthese van het huidpigment melanine en bevordert een toename van de grootte en het aantal pigmentcellen (melanocyten) in de huid van amfibieën. Deze eigenschappen van MSH worden gebruikt bij biologische testen van het hormoon. Er zijn twee soorten van het hormoon: alfa-MSH en bèta-MSH. Het is aangetoond dat alfa-MSH niet soortspecifiek is en dezelfde chemische structuur heeft bij alle zoogdieren. Het molecuul is een peptideketen bestaande uit 13 aminozuren. Bèta-MSH daarentegen is wel soortspecifiek en de structuur verschilt per diersoort. Bij de meeste zoogdieren bestaat het bèta-MSH-molecuul uit 18 aminozuren, en alleen bij mensen is het vanaf het aminozuuruiteinde verlengd met vier aminozuren. Opgemerkt moet worden dat alfa-MSH enige adrenocorticotrope activiteit heeft en dat het effect ervan op het gedrag van dieren en mensen inmiddels is bewezen.
Oxytocine en vasopressine
Vasopressine en oxytocine, die in de hypothalamus worden gesynthetiseerd, hopen zich op in de achterkwab van de hypofyse: vasopressine in de neuronen van de nucleus supraopticus en oxytocine in de nucleus paraventriculatoire. Vervolgens worden ze naar de hypofyse getransporteerd. Benadrukt moet worden dat de voorloper van het vasopressinehormoon eerst in de hypothalamus wordt gesynthetiseerd. Tegelijkertijd worden daar de eiwitten neurofysine type 1 en 2 geproduceerd. De eerste bindt oxytocine en de tweede bindt vasopressine. Deze complexen migreren in de vorm van neurosecretoire granula in het cytoplasma langs het axon en bereiken de achterkwab van de hypofyse, waar de zenuwvezels in de vaatwand eindigen en de inhoud van de granula in het bloed terechtkomt. Vasopressine en oxytocine zijn de eerste hypofysehormonen met een volledig vastgelegde aminozuurvolgorde. In hun chemische structuur zijn het nonapeptiden met één disulfidebrug.
De onderzochte hormonen hebben verschillende biologische effecten: ze stimuleren het transport van water en zouten door de membranen, hebben een vasopressieve werking, versterken de samentrekkingen van de gladde spieren van de baarmoeder tijdens de bevalling en verhogen de secretie van de borstklieren. Vasopressine heeft een sterkere antidiuretische werking dan oxytocine, terwijl oxytocine een sterker effect heeft op de baarmoeder en de borstklier. De belangrijkste regulator van de vasopressinesecretie is waterconsumptie; in de niertubuli bindt het zich aan receptoren in de cytoplasmatische membranen, met daaropvolgende activering van het enzym adenylaatcyclase in deze receptoren. Verschillende delen van het molecuul zijn verantwoordelijk voor de binding van het hormoon aan de receptor en voor het biologische effect.
De hypofyse, die via de hypothalamus verbonden is met het gehele zenuwstelsel, verenigt het endocriene systeem tot een functioneel geheel en draagt bij aan de stabilisatie van de interne omgeving van het lichaam (homeostase). Binnen het endocriene systeem vindt homeostatische regulatie plaats op basis van het principe van feedback tussen de voorkwab van de hypofyse en de doelklieren (schildklier, bijnierschors, geslachtsklieren). Een teveel aan het door de doelklier geproduceerde hormoon remt, en een tekort stimuleert de secretie en afgifte van het corresponderende tropische hormoon. De hypothalamus maakt deel uit van het feedbacksysteem. Hierin bevinden zich de receptorzones die gevoelig zijn voor de hormonen van de doelklieren. Door zich specifiek te binden aan hormonen die in het bloed circuleren en de respons te veranderen afhankelijk van de hormoonconcentratie, geven hypothalamische receptoren hun effect door aan de corresponderende hypothalamische centra, die het werk van de adenohypofyse coördineren en adenohypofysiotrope hormonen van de hypothalamus afgeven. De hypothalamus moet daarom worden beschouwd als een neuro-endocrien brein.